エネルギー交換-これは複雑な有機化合物の段階的な分解であり、エネルギーの放出を進めます。エネルギーはATP分子のマクロ作動性結合に保存され、生合成を含む細胞の生命の過程で使用されます。 プラスチック交換。
好気性生物は以下を生成します:
- 準備-生体高分子のモノマーへの分割。
- 無酸素解糖は、ブドウ糖のピルビン酸への分解です。
- 空気-ピルビン酸を二酸化炭素と水に分解します。
準備段階
エネルギー代謝の準備段階で、食物とともに受け取った有機化合物は、より単純な、通常はモノマーに分解されます。 したがって、炭水化物はブドウ糖を含む糖に分解されます。 タンパク質-アミノ酸へ; 脂肪-グリセロールと脂肪酸に。
エネルギーは放出されますが、ATPに蓄えられないため、後で使用することはできません。 エネルギーは熱の形で放散されます。
多細胞複合動物のポリマーの分解は、ここで腺から分泌される酵素の作用下で消化管で進行します。 次に、形成されたモノマーは主に腸を介して血液に吸収されます。 栄養素は血球を介して輸送されます。
ただし、すべての物質が消化器系でモノマーに分解するわけではありません。 多くの分裂は、細胞内のリソソームで直接起こります。 単細胞生物では、吸収された物質は消化液胞に入り、そこで消化されます。
得られたモノマーは、エネルギー交換とプラスチック交換の両方に使用できます。 前者の場合、それらは分割され、後者の場合、セル自体のコンポーネントはそれらから合成されます。
エネルギー代謝の無酸素段階
無酸素段階は細胞の細胞質で起こり、好気性生物の場合は、 解糖系-グルコースの酵素的多段階酸化とピルビン酸への分解、ピルビン酸とも呼ばれます。
グルコース分子には6つの炭素原子が含まれています。 解糖の間に、それは3つの炭素原子を含むピルビン酸の2つの分子に分解されます。 この場合、水素原子の一部が分離され、NAD補酵素に移動し、NAD補酵素が酸素段階に関与します。
解糖中に放出されるエネルギーの一部はATP分子に蓄えられます。 グルコース1分子あたり2つのATP分子のみが合成されます。
NADに貯蔵されているピルビン酸に残っているエネルギーは、エネルギー代謝の次の段階で好気性菌からさらに抽出されます。
嫌気性条件下では、細胞呼吸の酸素段階がない場合、ピルビン酸は乳酸に「中和」されるか、発酵します。 この場合、エネルギーは蓄えられません。 したがって、ここでは、有用なエネルギー出力は、非効率的な解糖によってのみ提供されます。
酸素段階
酸素ステップはミトコンドリアで起こります。 クレブス回路と酸化的リン酸化の2つのサブステージがあります。 セルに入る酸素は2番目にのみ使用されます。 クレブス回路は二酸化炭素を生成して放出します。
クレブス回路ミトコンドリアのマトリックスで起こり、多くの酵素によって実行されます。 ピルビン酸分子自体(または酪酸、アミノ酸)を受け取りませんが、前者のピルビン酸の2つの炭素原子を含む補酵素Aの助けを借りてアセチル基を分離します。 多段階のクレブス回路の間に、アセチル基は2つのCO2分子と水素原子に分割されます。 水素はNADおよびFADと結合します。 GDP分子の合成も起こり、後でATPの合成につながります。
1つのグルコース分子に対して2つのクレブス回路があり、そこから2つのピルビン酸が形成されます。 したがって、2つのATP分子が形成されます。 エネルギー代謝がここで終了した場合、グルコース分子の完全な分解により、4つのATP分子(解糖系から2つ)が得られます。
酸化的リン酸化クリステで発生します-ミトコンドリアの内膜の成長。 それは酵素と補酵素のコンベヤーによって提供され、いわゆる呼吸鎖を形成し、酵素ATPシンテターゼで終わります。
水素と電子は、補酵素NADとFADから呼吸鎖を介して伝達されます。 転送は、水素プロトンがミトコンドリア内膜の外側に蓄積し、連鎖の最後の酵素が電子のみを転送するように実行されます。
最終的に、電子は膜の内側にある酸素分子に移動し、その結果、電子は負に帯電します。 電位勾配の臨界レベルが発生し、ATPシンテターゼのチャネルを介したプロトンの移動につながります。 水素プロトンの移動エネルギーはATP分子の合成に使用され、プロトン自体が酸素アニオンと結合して水分子を形成します。
ATP分子で表される呼吸鎖の機能のエネルギー出力は大きく、1つの初期グルコース分子あたり合計で32から34のATP分子の範囲です。
すべての体は正しく機能するためにエネルギーを必要とします。 人は代謝を通してそれを受け取ります、それは外部からの受け取りを条件として可能です。 必要量タンパク質、脂肪、炭水化物。 このプロセスは常に発生します。 受け取ったエネルギーと消費したエネルギーのバランスが崩れなければ、代謝は正常です。 その失敗は、気分のむらから病院のベッドまで、健康状態の悪化につながる可能性があります。
代謝が妨げられる理由
代謝不良には多くの理由があります。 主なものを見つけるには、あなたのライフスタイルを分析する必要があります:
- 栄養は定期的でバランスが取れている必要があります。
- 睡眠-強くていっぱい;
- 動き-定期的かつアクティブ;
- 空気は新鮮できれいです。
- 気分-良い;
- ビタミンと微量元素の完全なセット。
スポーツをする人は、ダイエットに従う必要性とその利点について知っています 新鮮な空気。 これが彼らの生き方です。 国会にも存在する権利があります。 しかし、消費された製品の品質はしばしば基準を満たしていません。 また、ボリュームは必ずしも正確に計算できるとは限りません。 しかし、人間の臓器が正常に機能するために必要な有用な要素の主な供給源は食品です。 不十分で、タイムリーでなく、バランスの取れていない栄養のために、代謝過程で失敗が起こります。
ビタミンやミネラルは何のためにありますか?
残念ながら、人体はビタミンを生成することができません。 それらの主な機能は、代謝を調節し、さまざまなプロセスの正常な経過を確保することです。 造血、心血管、神経および 消化器系、酵素形成、有害作用への耐性 環境-これはすべて提供されます 通常レベル体内のビタミン。 それらのそれぞれは彼の領域に責任があります。
ビタミンのように、微量元素 化学物質)体が少量必要としますが、それらの不足はすべての重要なシステムの機能に大きく影響します。 それらは絶えず体から排泄されるので、それらの定期的な補充が必要です。
ビタミンと微量元素の蓄えを補充する方法
人間の生活では、栄養素の需要が高まる特別な時期があります。 季節の脚気を除外すると、これは成長と大きな運動の時間です(つまり、子供とアスリートにとってすべてが最高です)。 高品質の製品を自然に使用することで、在庫を補充できるとは限りません。 大手製薬会社によって特別に開発された製剤が助けになります。 それで、50年以上の間、家族のアメリカの会社NOW Foodsはそれらを生産してきました:自然、有機、健康、すなわち、自然、有機、健康。
神経系や心臓血管系に問題がある場合、免疫力が低下して内分泌系が破壊されたときは、薬に注意を払う必要があります 錠剤でビタミンB6。
サプリメントは薬ではなく、病気を予防したり、治癒プロセスをスピードアップしたりするのに役立ちます。 したがって、病気を待たないでください。 製品に十分な量のビタミンが供給されていないと感じた場合は、予防コースを飲むことができます。
ビタミンB群の利点は何ですか
体が完全に機能するためのこれらの要素の重要性を過大評価することは困難です。
チアミン(B1)食物の吸収に好影響を与え、すべてのシステムの働きを正常化します。
リボフラビン(B2)すべてに役立ちます 代謝過程優れた抗酸化剤。
ニコチン(B3)主に血管に影響を与えます。
シアノコバラミン(B12)腸で合成することができ、脂肪と炭水化物の代謝を調節します。 それは正常な成長の最も重要な要因の1つであり、神経障害を防ぐのに役立ち、男性の生殖能力に責任があります。
ビタミンB6(ピリドキシン)-このグループで最も人気のあるものの1つ。理由は次のとおりです。
- タンパク質の代謝と吸収に積極的に関与し、筋肉量の増加を助けます。
- 血中のコレステロールと脂質のレベルを低下させます。
- 心筋の働きを改善します。
- セロトニンの生成に関与するため、神経系に有益な効果があります。
- 肝臓を正常化します。
- 抗酸化剤として作用し、老化プロセスを遅らせます。
- けいれんや筋肉のけいれんを軽減します。
ひどい 身体活動ビタミンB6の基準は2倍にする必要があります。 この場合、在庫は人為的に補充するのが最も簡単です。 複雑な準備にも含まれています。
たとえば、Now Foodsは、B6に加えて、すべての人間のシステムに有益な効果を持つマグネシウムと亜鉛を含むZMAビタミン複合体を生成します。 この栄養補助食品は、アスリートの体の要素の不足を補うために特別に設計されています。 マグネシウムは、テストステロンのレベルを変えることによって筋力を高めるのに役立ちます。 残念ながら、この物質の十分な量は食品に由来していません。 そして、その欠乏はタンパク質の形成を阻害し、脳のプロセスを遅くし、仕事の混乱を引き起こします。 神経系。 結果として:
- ふくらはぎの筋肉のけいれんやけいれんが現れます。
- 圧力が上昇します。
- 心臓のリズムが乱れます。
- 倦怠感とうつ病が現れます。
マグネシウムの量の変化は、筋肉内のアミノ酸の形成、テストステロンおよび成長ホルモンの生成に関与する亜鉛の量の減少につながります。 彼から 十分でない免疫システムと性ホルモンの合成が苦しんでいます。 それは脂肪分解のレベルを高め、肝臓の問題を防ぎます。
栄養補助食品のすべての成分は完全に相互作用し、人体へのより効果的な効果に貢献します。 ZMA複合体は、筋肉量を構築するための優れたツールです。
ビタミンとミネラルのバランスを維持する方法
アスリートの膨大な負荷が体の衰弱につながることがあります。 この理由は、すべての人間のシステムの正常な機能に必要な物質のバランスを乱す栄養失調、ストレスおよび他の要因である可能性があります。 したがって、ビタミン複合体が 子供時代、およびそれらの正しいアプリケーションは、肯定的な結果のみを提供します。 より高い用量を服用することが達成に役立つとは思わないでください 最高の効果。 過剰なビタミンやミネラルは、 否定的な結果したがって、製造業者は徹底的な調査を実施し、最適な用量を計算します。
NOWFoodsオンラインストアが提供する栄養補助食品は薬ではありません。 身体の働きにおける無視された形の障害の治療を頼りにする価値はありません。 生物学的に活性な複合体は、回復プロセスをスピードアップしたり、病気を予防したりするための優れた方法です。 それらはすべてヘルスケアを念頭に置いて作成されています。
エネルギーはどの程度正確に蓄えられていますか ATP(アデノシン三リン酸)、そしていくつかの有用な仕事をすることはどのように与えられますか? ある抽象的なエネルギーが生細胞内にある分子の形で物質担体を突然受け取り、それが熱の形ではなく(多かれ少なかれ明確である)、次の形で放出されることは非常に複雑に思えます。別の分子を作成します。 通常、教科書の著者は、「エネルギーは分子の部分間の高エネルギー結合の形で蓄積され、この結合が切断されると与えられ、有用な仕事をする」というフレーズに限定されますが、これは何も説明していません。

最も一般的な用語では、分子とエネルギーによるこれらの操作は次のように発生します。 または、それらは同様の反応の連鎖で葉緑体で作成されます。 これは、ミトコンドリア内の栄養素の制御された燃焼から得られたエネルギーまたは光子のエネルギーを浪費します 日光クロロフィル分子に落ちる。 次に、ATPは、いくつかの作業を行う必要があるセル内の場所に配信されます。 そして、1つまたは2つのリン酸基がそこから切断されると、エネルギーが放出され、これがこの働きをします。 同時に、ATPは2つの分子に分解されます。1つのリン酸基のみが切断されると、ATPは次のようになります。 ADP(アデノシン二リン酸。これは、非常に分離したリン酸基がない場合にのみアデノシン三リン酸と異なります)。 ATPが一度に2つのリン酸基を放棄した場合、より多くのエネルギーが放出され、アデノシンMONOリン酸がATPから残ります( AMF).
明らかに、細胞は逆のプロセスを実行する必要があり、ADPまたはAMP分子をATPに変換して、サイクルを繰り返すことができます。 しかし、これらの「ブランク」分子は、ATPへの変換に欠けているリン酸塩の隣を簡単に泳ぐことができ、そのような会合反応はエネルギー的に不利であるため、それらと結合することはありません。

化学反応の「エネルギーの利点」とは何かを知っていれば理解するのは非常に簡単です 熱力学の第二法則:宇宙や他のシステムから隔離されたシステムでは、無秩序は成長することしかできません。 つまり、この法則に従って、細胞内に整然と並んでいる複雑に組織化された分子は、破壊され、より小さな分子を形成するか、個々の原子に分解することさえできます。 この考えを理解するために、複雑な分子をレゴから組み立てられた飛行機と比較することができます。 次に、複雑な分子が分裂する小分子は、この航空機の個々の部分に関連付けられ、原子は個々のレゴブロックに関連付けられます。 きちんと組み立てられた平面を見て、それを部品の寄せ集めと比較すると、複雑な分子が小さな分子よりも多くの秩序を含んでいる理由が明らかになります。
このような(航空機ではなく分子の)崩壊反応はエネルギー的に有利であり、これは自発的に実行できることを意味し、崩壊中にエネルギーが放出されます。 実際には、航空機の分割はエネルギー的に有益ですが、パーツ自体が互いに分割されず、外力がこれらを使用したい子供の形でそれらの分離をパフする必要があるという事実にもかかわらず他の何かのための部品、彼は飛行機を部品の混沌とした山に変えるために高度に注文された食べ物を食べることから得られたエネルギーを費やします。 また、パーツがしっかりとくっついているほど、熱の形で放出されるなど、より多くのエネルギーが消費されます。 結論:パン(エネルギー源)の一部と飛行機が混沌とした塊に変わり、子供の周りの空気分子が加熱されます(したがって、よりランダムに移動します)-より多くの混乱があります、つまり、飛行機を分割することはエネルギー的にです有益。
要約すると、熱力学の第二法則に従って、次のルールを定式化できます。
1.秩序の量が減少すると、エネルギーが放出され、エネルギー的に好ましい反応が起こります
2.注文量が増えると、エネルギーが吸収され、エネルギーを消費する反応が起こります
一見、秩序から混沌へのこの避けられない動きは、単一の受精卵と母牛によって吸収された栄養分子から構築するなどのプロセスを逆転させることを不可能にします。
しかし、それでも、これは起こります。その理由は、生物には、エントロピーに対する宇宙の欲求をサポートし、自分自身とその子孫を構築することを可能にする1つの機能があるためです。 2つの反応を1つのプロセスに結合し、一方はエネルギー的に有利であり、もう一方はエネルギー集約的です。 このような2つの反応の組み合わせにより、最初の反応中に放出されるエネルギーが2番目の反応のエネルギーコストを超えることを保証できます。 飛行機の例では、飛行機を別々に分解するのはエネルギーを消費し、男の子の新陳代謝によって破壊されたパンの形の外部エネルギー源がなければ、飛行機は永遠に立ちます。

それはそりで下り坂を行くようなものです。まず、人は食べ物を食べながら、高度に秩序化された鶏肉を体内の分子と原子に分割するエネルギー的に有利なプロセスの結果として得られたエネルギーを蓄えます。 そして、彼はこのエネルギーを使って、そりを山に引き上げます。 そりを下から上に動かすことはエネルギー的に不利であるため、そりが自然に転がることはありません。これには、何らかのサードパーティのエネルギーが必要です。 そして、鶏肉を食べることで得られるエネルギーが登りを克服するのに十分でない場合、「そりで山を転がる」プロセスは起こりません。
それはエネルギーを消費する反応です( エネルギーを消費する反応 )カップリング反応で放出されるエネルギーを吸収することにより、秩序の量を増やします。 そして、これらのカップリング反応におけるエネルギーの放出と消費のバランスは常に正でなければなりません。つまり、それらの組み合わせはカオスの量を増やします。 増加の例 エントロピ(障害) ( エントロピ[‘entrəpɪ])は、エネルギーを与える反応中の熱の放出です( エネルギー供給反応):反応した分子に隣接する物質の粒子は、反応する分子からエネルギーショックを受け、より速く、より無秩序に動き始め、この物質と隣接する物質の他の分子と原子を順番に押します。

再び食べ物からエネルギーを得ることに戻ります。バノフィーパイは、胃に入った咀嚼塊よりもはるかに注文が多いです。 これは、腸がそれを分割する分子よりも大きく、より秩序だった分子で構成されています。 そして、それらは順番に、体の細胞に運ばれ、そこで個々の原子や電子さえもそれらから引き裂かれます...そして、一枚のケーキの混沌の増加の各段階で、エネルギーは解放され、幸せな食べる人の器官とオルガネラが捕らえ、ATP(エネルギー消費)の形でそれを保存し、新しい必要な分子の構築(エネルギー消費)または体の加熱(エネルギー消費)を可能にします消費する)。 その結果、「人-バノフィーパイ-宇宙」システムの秩序は少なくなりますが(ケーキの破壊とそれを処理する器官による熱エネルギーの放出のため)、単一の人体では、幸福はより秩序になる(新しい分子、オルガネラの一部、細胞器官全体の出現による)。
ATP分子に戻ると、この熱力学的余談の後、エネルギー的に有利な反応から受け取ったエネルギーを、その構成部分(より小さな分子)から作成するために費やす必要があることが明らかになります。 それを作成する1つの方法が詳細に説明されており、別の(非常に類似した)葉緑体で使用され、プロトン勾配のエネルギーの代わりに、太陽から放出される光子のエネルギーが使用されます。

ATPを生成する反応には3つのグループがあります(右の図を参照)。
- ブドウ糖と酪酸が細胞質内で大きな分子に分解されることで、すでに一定量のATPを得ることができます(小さな、この段階で分割された1つのブドウ糖分子に対して2つのATP分子しか得られません)。 しかし、この段階の主な目的は、ミトコンドリアの呼吸鎖で使用される分子を作成することです。
- ミトコンドリアマトリックスで発生するクレブス回路の前の段階で得られた分子をさらに分割すると、ATP分子が1つだけ得られます。その主な目標は、前の段落と同じです。
- 最後に、前の段階で蓄積された分子は、ATPの生成のためにミトコンドリアの呼吸鎖で使用され、ここでその多くが放出されます(これについては以下で詳しく説明します)。
これらすべてをより詳細に説明し、エネルギーの生産と消費の観点から同じ反応を見ると、次のようになります。
0. 食物分子は、細胞の細胞質で発生する一次切断、およびミトコンドリアマトリックスですでに発生している「クレブス回路」と呼ばれる一連の化学反応で注意深く燃焼(酸化)されます- エネルギーを生み出す準備段階の一部。
他のエネルギー的に有利な反応との共役の結果として、新しい分子、2つのATP分子および他の物質のいくつかの分子を作成するというすでにエネルギー的に不利な反応が形成されます- エネルギーを消費する準備段階の一部。 これらの共形成分子は、次の段階でミトコンドリア呼吸鎖で使用される高エネルギー電子のキャリアです。

1. ミトコンドリア、バクテリア、およびいくつかの古細菌の膜では、前の段階で得られた分子(ATPからではない)からのプロトンと電子のエネルギーを与える分裂が起こります。 呼吸鎖の複合体(左の図のI、III、IV)を通る電子の通過は黄色の曲がりくねった矢印で示され、プロトンのこれらの複合体(したがってミトコンドリア内膜)を通る通過は次のように示されます。赤い矢印。
強力な酸化剤である酸素を使用して、電子をキャリア分子から単純に分離できないのはなぜですか。放出されたエネルギーを使用できます。 最終的に同じ酸素になるので、なぜそれらをある複合体から別の複合体に移すのですか? 電子を与えることで電子を引き付ける能力の違いが大きいことがわかります( 還元剤)および電子収集( 酸化剤)電子移動反応に関与する分子ほど、この反応中に放出されるエネルギーが多くなります。
クレブス回路で形成される電子と酸素のキャリア分子のこの能力の違いは、この場合に放出されるエネルギーがいくつかのATP分子の合成に十分であるようなものです。 しかし、システムのエネルギーがこのように急激に低下するため、この反応はほとんど爆発的な力で進行し、ほとんどすべてのエネルギーが捕捉されていない熱の形で放出されます。つまり、実際には無駄になります。

一方、生細胞はこの反応をいくつかの小さな段階に分割し、最初に電子を弱い引力のキャリア分子から呼吸鎖のわずかに強い引力の最初の複合体に、それからさらにわずかに強い引力に転送します ユビキノン(また コエンザイムQ-10)、そのタスクは、電子を次の、さらにわずかに強く引き付ける呼吸複合体に引きずり込むことです。呼吸複合体は、この失敗した爆発からエネルギーの一部を受け取り、膜を通してプロトンをポンプで送ります。 、それに引き付けられ、いくつかのプロトンをつかみ、水分子を形成しません。 1つの強力な反応を小さなステップに分割することで、有用なエネルギーのほぼ半分を有用な仕事の実行に向けることができます。この場合は、 プロトン電気化学勾配これについては、2番目の段落で説明します。
転送された電子のエネルギーが、膜を介してプロトンをポンピングするエネルギーを消費する結合反応をどの程度正確に助けるかは、まだ解明され始めたばかりです。 おそらく、帯電した粒子(電子)の存在は、それが配置されている膜に埋め込まれたタンパク質内の場所の構成に影響を与えます。そのため、この変化により、プロトンがタンパク質に引き込まれ、タンパク質チャネルを通って移動します。膜に。 実際、高エネルギー電子がキャリア分子から分裂し、最終的に酸素に移動した結果として得られるエネルギーが、プロトン勾配の形で保存されることが重要です。
2. 膜の外側のポイント1からのイベントの結果として蓄積され、内側に到達する傾向があるプロトンのエネルギーは、2つの一方向の力で構成されます。
- 電気(プロトンの正電荷は膜の反対側の負電荷の蓄積の場所に行く傾向があります)そして
- 化学(他の物質の場合と同様に、陽子は空間に均一に分散しようとし、陽子の濃度が高い場所から少ない場所に広がります)

内膜の負に帯電した側へのプロトンの電気的引力は、プロトン濃度の違いのためにプロトンが低濃度の場所に移動する傾向よりもはるかに強力です(これは、上の図)。 これらの駆動力の合計エネルギーは非常に大きいため、プロトンを膜内で移動させ、付随するエネルギー消費反応、つまりADPとリン酸塩からのATPの生成を促進するのに十分です。
これがなぜエネルギーを必要とするのか、そしてプロトン吸引のエネルギーがATP分子の2つの部分の間の化学結合のエネルギーにどのように正確に変換されるのかをより詳細に考えてみましょう。

ADP分子(図の右側)は、別のリン酸基を獲得することを望んでいません。この基が結合できる酸素原子は、リン酸と同じくらい負に帯電しています。つまり、互いに反発します。 一般に、ADPは反応せず、化学的に受動的です。 次に、リン酸はそのリン原子に結合した独自の酸素原子を持っています。これは、ATP分子を作成するときにリン酸とADPの間の結合のサイトになる可能性があるため、主導権を握ることもできません。
したがって、これらの分子は1つの酵素によって接続され、展開されて、それらと「余分な」原子との間の結合が弱まり、壊れてから、これらの分子の2つの化学的に活性な末端をもたらし、原子が不足および過剰になる必要があります。電子、お互いに。
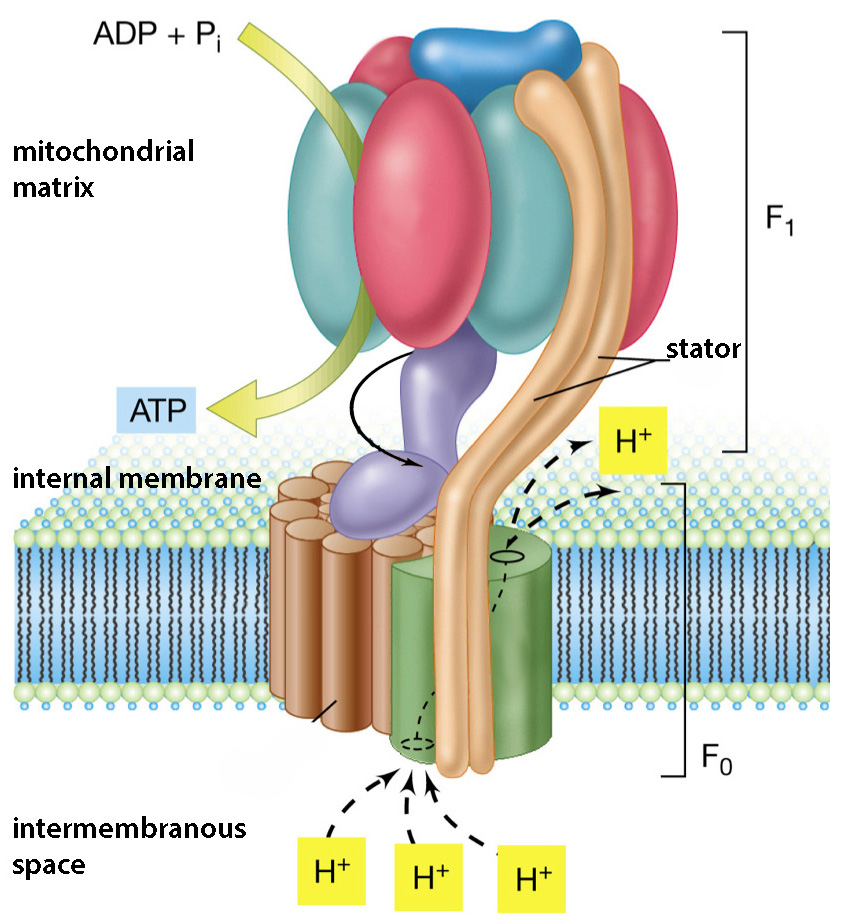
相互に到達する領域に落ちたリン(P +)と酸素(O-)のイオンは、元々酸素に属していた1つの電子を共同で所有しているため、強力な共有結合によって結合されます。 この分子処理酵素は ATP合成酵素、およびその構成と 相対位置それはそれを通過するプロトンからADPとリン酸塩を受け取ります。 プロトンが膜の反対に帯電した側に到達することはエネルギー的に有利であり、さらに、それらはほとんどなく、 唯一の方法プロトンが同時に回転する「ローター」である酵素を通過します。
ATP合成酵素の構造を右の図に示します。 陽子の通過によるその回転要素が強調表示されます 紫の、および下の動画は、その回転とATP分子の作成の図を示しています。 酵素はほとんど分子モーターのように働き、回転します 電気化学陽子の現在のエネルギー 力学的エネルギー 2セットのタンパク質の相互摩擦:回転する「脚」が「マッシュルームキャップ」の不動のタンパク質をこすり、「キャップ」のサブユニットがその形状を変化させます。 この機械的変形は 化学結合エネルギー ATPの合成において、ADPとリン酸分子が処理され、それらの間の共有結合の形成に必要な方法で展開される場合。

各ATPシンターゼは、毎秒最大100個のATP分子を合成することができ、合成されたATP分子ごとに、約3つのプロトンがシンテターゼを通過する必要があります。 細胞内で合成されるATPのほとんどはこのように形成されており、ミトコンドリアの外側で発生する食物分子の一次処理の結果であるのはごく一部です。
いつでも、典型的な生細胞には約10億のATP分子があります。 多くのセルでは、このATPはすべて1〜2分ごとに交換(つまり、使用および再作成)されます。 安静時の平均的な人は、24時間ごとに自分の質量とほぼ等しいATPの質量を使用します。

一般に、グルコースまたは酪酸の二酸化炭素と水への酸化中に放出されるエネルギーのほぼ半分が捕捉され、ADPとリン酸からのATP形成のエネルギー的に不利な反応に使用されます。 50%の効率は悪くありません。たとえば、自動車のエンジンは、燃料に含まれるエネルギーの20%だけを有用な仕事に使用します。 同時に、どちらの場合も残りのエネルギーは熱の形で放散され、一部の車と同様に、動物は常にこの過剰を(もちろん完全ではありませんが)体の温めに費やします。 ここで述べた反応の過程で、1つのグルコース分子が徐々に二酸化炭素と水に分解され、細胞に30個のATP分子を供給します。
したがって、エネルギーがどこから来て、ATPにどれだけ正確に保存されているかによって、すべてが多かれ少なかれ明確になります。 理解することは残っています 蓄積されたエネルギーがどのように正確に与えられるか、そしてこの場合何が起こるか分子原子レベルで。
ADPとリン酸の間に形成される共有結合は 高エネルギー 2つの理由:
- それが故障すると、それは多くのエネルギーを放出します。
- この結合の作成に関与する電子(つまり、この結合が形成される酸素原子とリン原子の周りを回転する)は高エネルギーです。つまり、原子核の周りの「高」軌道にあります。 そして、彼らがより低いレベルにジャンプして過剰なエネルギーを放出することはエネルギー的に有益ですが、彼らがこの場所にいて、酸素とリンの原子を固定している限り、彼らは「ジャンプ」することができません。
電子がより便利な低エネルギー軌道に落ちるというこの欲求は、高エネルギー結合の破壊の容易さと、光子(電磁相互作用のキャリア)の形で放出されるエネルギーの両方を保証します。 どの分子が崩壊するATP分子の酵素によって置換されるかに応じて、どの分子が電子によって放出された光子を吸収しますか? さまざまなバリエーションイベント。 しかし毎回 高エネルギー結合の形で蓄積されたエネルギーは、セルのいくつかのニーズに使用されます:
シナリオ1:リン酸塩は別の物質の分子に移すことができます。 この場合、高エネルギー電子は、すでにリン酸とこのレシピエント分子の極限原子との間に新しい結合を形成します。 このような反応が起こるための条件は、そのエネルギー的利点です。この新しい結合では、電子はATP分子の一部であったときよりもわずかに少ないエネルギーを持ち、エネルギーの一部を光子の形で外側に放出する必要があります。

このような反応の目的は、レシピエント分子を活性化することです(左の図では、それが示されています) で-OH):リン酸塩を添加する前は、それは受動的であり、別の受動的分子と反応することができませんでした しかし、しかし今、彼女は高エネルギー電子の形でエネルギーの蓄えの所有者です、それは彼女がそれをどこかで使うことができることを意味します。 たとえば、分子をそれ自体に結合する しかし、そのような耳のフェイント(つまり、結合電子の高エネルギー)がなければ、取り付けることはできません。 その後、リン酸塩はその仕事を終えて切り離されます。
これにより、一連の反応が発生します。
1. ATP+パッシブ分子 で ➡️ ADP+結合したリン酸塩による活性分子 V-R
2. 活性化された分子 V-R+パッシブ分子 しかし➡️接続された分子 A-B+リン酸塩を分離する( R)
これらの反応は両方ともエネルギー的に有利です。それぞれが高エネルギーの結合電子を含み、1つの結合が切断されて別の結合が形成されると、そのエネルギーの一部が光子放出の形で失われます。 これらの反応の結果として、2つの受動分子が接続されます。 これらの分子を直接つなぐ反応を考えると(受動分子 で+パッシブ分子 しかし➡️接続された分子 A-B)、それからそれはエネルギー的に高価であることが判明し、行うことができません。 細胞は、この反応を、上記の2つの反応中に、エネルギー的に有利なATPのADPとリン酸への分裂と組み合わせることによって「不可能なことをします」。 分裂は2つの段階で発生し、結合電子のエネルギーの各部分が有用な仕事を行うために費やされます。つまり、2つの分子間に必要な結合を作成し、そこから3番目の分子を取得します( A-B)細胞の機能に必要です。
シナリオ2:リン酸はATP分子から同時に分離することができ、放出されたエネルギーは酵素または作用するタンパク質によって捕捉され、有用な仕事をするために費やされます。

取るに足らない憤りのように知覚できないものをどうやって捕まえることができますか 電磁界電子がより低い軌道に落ちるとき? 非常に簡単です。他の電子の助けを借りて、そして電子によって放出された光子を吸収することができる原子の助けを借りて。

分子を構成する原子は、によって強力な鎖と環にまとめられています(このような鎖は右の写真の折りたたまれていないタンパク質です)。 そして、これらの分子の別々の部分は、より弱い電磁相互作用(たとえば、水素結合やファンデルワールス力)によって互いに引き付けられ、複雑な構造を形成することができます。 原子のこれらの構成のいくつかは非常に安定しており、電磁場の乱れがそれらを揺さぶることはありません..揺れません..一般的に、それらは安定しています。 そして、いくつかは非常に可動性があり、わずかな電磁キックで構成を変更するのに十分です(通常、これらは共有結合ではありません)。 そして、まさにそのようなキックは、リン酸塩が分離されたときに下の軌道に渡された電子によって放出された、電磁場の非常に到着した光子キャリアによって彼らに与えられます。
ATP分子の分解の結果としてのタンパク質の構成の変化は、細胞内で発生する最も驚くべきイベントの原因です。 確かに、少なくとも「YouTubeでアニメーションを見る」レベルで細胞プロセスに興味を持っている人は、タンパク質分子を示すビデオに出くわしました。 キネシン、文字通り歩いて、足を再配置し、細胞の骨格の糸に沿って、それに付けられた荷物を引きずります。
このステッピングを提供するのはATPからのリン酸塩の分割であり、その方法は次のとおりです。

キネシン( キネシン)自発的に変化する傾向がある特殊なタイプのタンパク質を指します コンフォメーション(分子内の原子の相互位置)。 そのままにしておくと、1つの「脚」でアクチンフィラメントに結合しているコンフォメーション1からランダムに遷移します( アクチンフィラメント)-最も細いねじ山形成 細胞骨格セル( 細胞骨格)、コンフォメーション2になり、一歩前進して2つの「脚」の上に立ちます。 コンフォメーション2から、コンフォメーション3(後脚を前脚に取り付ける)とコンフォメーション1の両方に等しい確率で通過します。したがって、キネシンはどの方向にも移動せず、単に目的もなくさまようだけです。

しかし、ATP分子と結合するとすぐにすべてが変化します。 左の図に示すように、コンフォメーション1のキネシンにATPを加えると、空間位置が変化し、コンフォメーション2に移行します。これは、ATPとキネシンの相互の電磁的影響によるものです。お互いの分子。 エネルギーが消費されていないため、この反応は可逆的であり、ATPがキネシンから分離すると、ATPは単にその「脚」を持ち上げ、所定の位置に留まり、次のATP分子を待ちます。
しかし、それが長引くと、これらの分子の相互引力により、ATP内にリン酸を保持している結合が破壊されます。 同時に放出されるエネルギー、およびATPの2つの分子への分解(電磁場によってキネシン原子にすでに異なる影響を与える)は、キネシンのコンフォメーションが変化するという事実につながります。 」。 ADPとリン酸が分離し、キネシンが元のコンフォメーションに戻るときに起こる一歩を踏み出す必要があります1。
ATP加水分解の結果、キネシンは右に移動し、次の分子がそれに加わるとすぐに、そこに蓄えられたエネルギーを使用して、さらにいくつかのステップを踏みます。
ADPとリン酸が結合したコンフォメーション3のキネシンは、「バックステップ」を実行してもコンフォメーション2に戻れないことが重要です。 これは、体温調節の第2法則に準拠するという同じ原則によって説明されます。つまり、「キネシン+ ATP」システムのコンフォメーション2からコンフォメーション3への移行にはエネルギーの放出が伴います。つまり、逆の移行はエネルギーになります。消費する。 それが起こるためには、ADPをリン酸塩と組み合わせるためにどこかからエネルギーを奪う必要があり、この状況ではそれを奪う場所がありません。 したがって、ATPに接続されたキネシンは一方向にのみ開いており、セルの一方の端からもう一方の端に何かをドラッグすることで便利な作業を行うことができます。 たとえば、キネシンは、分裂中の細胞の染色体を引き離すことに関与しています。 有糸分裂(真核細胞を分裂させるプロセス)。 筋肉タンパク質 ミオシンアクチンフィラメントに沿って走り、筋肉の収縮を引き起こします。

この動きは非常に速いです:いくつか モーター(の責任者 様々な形態細胞の可動性)、遺伝子複製に関与するタンパク質は、毎秒数千ヌクレオチドの速度でDNA鎖に沿って突進します。
それらはすべて通過します 加水分解 ATP(水分子から取り出された原子が分解によって生じた小さな分子に追加された分子の破壊。加水分解はATPとADPの相互変換の図の右側に示されています)。 または加水分解によって GTP、ATPとは、別のヌクレオチド(グアニン)が含まれているという点でのみ異なります。
シナリオ3:ATPまたはヌクレオチドを含む別の同様の分子から一度に2つのリン酸基を除去すると、1つのリン酸のみを除去した場合よりもさらに多くのエネルギーが放出されます。 このような強力なリリースにより、DNAおよびRNA分子の強力な糖リン酸バックボーンを作成できます。

1. ヌクレオチドが構築中のDNAまたはRNA鎖に結合できるようにするには、2つのリン酸分子を結合してヌクレオチドを活性化する必要があります。 これは、細胞の酵素によって実行されるエネルギーを消費する反応です。
2. 酵素DNAまたはRNAポリメラーゼ(下の図には示されていません)は、活性化ヌクレオチド(GTPは図に示されています)を構築中のポリヌクレオチドに結合し、2つのリン酸基の切断を触媒します。 放出されたエネルギーは、あるヌクレオチドのリン酸基と別のヌクレオチドのリボースの間に結合を作成するために使用されます。 結果として作成される結合は高エネルギーではありません。つまり、それらは簡単に破壊されません。これは、細胞の遺伝情報を含むまたは伝達する分子を構築するための利点です。

自然界では、熱力学の第二法則により、エネルギー的に有利な反応のみが自発的に発生する可能性があります。
それにもかかわらず、生細胞は2つの反応を組み合わせることができ、そのうちの1つは、2つ目が吸収するよりも少し多くのエネルギーを与えるため、エネルギーを消費する反応を実行します。 エネルギーを消費する反応は、個々の分子や原子からより大きな分子、細胞オルガネラ、全細胞、組織、器官、多細胞生物を作り出すこと、そしてそれらの代謝のためにエネルギーを蓄えることを目的としています。
エネルギーの貯蔵は、有機分子の制御された段階的な破壊(エネルギー生成プロセス)と、エネルギー運搬分子の作成(エネルギー消費プロセス)によって実行されます。 光合成生物は、このようにしてクロロフィルによって捕らえられた太陽光子のエネルギーを蓄えます。
分子-エネルギーキャリアは2つのグループに分けられます:高エネルギー結合の形で、または付着した高エネルギー電子の形でエネルギーを貯蔵します。 ただし、最初のグループでは、同じ高エネルギー電子によって高エネルギーが提供されるため、エネルギーは駆動された状態で蓄積されていると言えます。 上級異なる分子の一部である電子
このようにして蓄えられたエネルギーは、高エネルギー結合を破壊する方法と、高エネルギー電子を転送して徐々にエネルギーを減らす方法の2つの方法で放出されます。 どちらの場合も、エネルギーは、電磁場(光子)と熱の粒子キャリアのより低いエネルギーレベルに通過する電子による放出の形で放出されます。 この光子は、有用な仕事が行われるように捕捉されます(最初のケースでは代謝に必要な分子の形成、2番目のケースではミトコンドリア膜を通してプロトンをポンピングします)
プロトン勾配の形で蓄えられたエネルギーは、ATPの合成だけでなく、この章の範囲を超えている他の細胞プロセスにも使用されます(そのサイズを考えると、誰も気分を害することはないと思います)。 そして、合成されたATPは、前の段落で説明したように使用されます。
太い木の豊富な成長、
不毛の砂に根ざしている
彼自身を承認し、明確に述べています
空気からの脂っこい脂肪の脂っこいシート
吸収する...
M.V.ロモノソフ
エネルギーはどのように細胞に蓄えられますか? 代謝とは何ですか? 解糖、発酵、細胞呼吸のプロセスの本質は何ですか? 光合成の明期と暗期ではどのようなプロセスが起こりますか? エネルギーとプラスチック交換のプロセスはどのように関連していますか? 化学合成とは何ですか?
レッスン-講義
ある種類のエネルギーを別の種類のエネルギーに変換する能力(放射エネルギーを化学結合のエネルギーに、化学エネルギーを機械的エネルギーになど)は、生物の基本的な特性の1つです。 ここでは、これらのプロセスが生物でどのように実現されているかを詳しく考察します。
ATP-細胞内のエネルギーの主なキャリア。 細胞の生命活動の兆候を実行するには、エネルギーが必要です。 独立栄養生物は光合成反応中に太陽から初期エネルギーを受け取りますが、従属栄養生物は食物からの有機化合物をエネルギー源として使用します。 エネルギーは細胞によって分子の化学結合に蓄えられます ATP(アデノシン三リン酸)、糖残基(リボース)と窒素塩基残基(アデニン)の3つのリン酸基からなるヌクレオチドです(図52)。
米。 52.ATP分子
リン酸残基間の結合は、それが切断されると、マクロ作動性と呼ばれます。 たくさんのエネルギー。 通常、細胞は末端のリン酸基のみを除去することによってATPからエネルギーを抽出します。 この場合、ADP(アデノシン二リン酸)、リン酸が形成され、40 kJ/molが放出されます。
ATP分子は細胞の普遍的なエネルギー交渉チップの役割を果たします。 それらは、有機化合物の酵素的合成、タンパク質の仕事(分子モーターまたは膜輸送タンパク質など)であるかどうかにかかわらず、エネルギー集約的なプロセスのサイトに配信されます。ATP分子の逆合成は、リン酸を結合することによって実行されますエネルギー吸収を伴うADPへのグループ。 細胞によるATPの形でのエネルギーの貯蔵は反応の間に実行されます エネルギー代謝。 彼は密接に関連しています プラスチック交換その間、細胞はその機能に必要な有機化合物を生成します。
細胞内の代謝とエネルギー(代謝)。 代謝-相互に関連する、プラスチックとエネルギー代謝のすべての反応の全体。 細胞内では、炭水化物、脂肪、タンパク質、核酸の合成が絶えず行われています。 化合物の合成には、常にエネルギーの消費が伴います。つまり、ATPの不可欠な参加が伴います。 ATPを形成するためのエネルギー源は、細胞に入るタンパク質、脂肪、炭水化物の酸化の酵素反応です。 このプロセスはエネルギーを放出し、ATPに蓄えられます。 グルコースの酸化は、細胞のエネルギー代謝において特別な役割を果たします。 グルコース分子は、一連の連続した変換を受けます。
と呼ばれる最初の段階 解糖、細胞の細胞質で起こり、酸素を必要としません。 酵素が関与する連続反応の結果として、グルコースはピルビン酸の2つの分子に分解されます。 この場合、2つのATP分子が消費され、酸化中に放出されるエネルギーは4つのATP分子を形成するのに十分です。 その結果、解糖のエネルギー収量は小さく、2つのATP分子になります。
C 6 H1 206→2C3H 4 0 3 + 4H + + 2ATP
嫌気性条件下(酸素がない場合)では、さらなる変換が関連する可能性があります さまざまなタイプ 発酵.
みんな知ってる 乳酸発酵(ミルクサワー)、乳酸菌やバクテリアの活動によって発生します。 解糖系とメカニズムが似ており、ここでの最終生成物は乳酸だけです。 このタイプのブドウ糖酸化は、勤勉な筋肉などの酸素欠乏細胞で起こります。 化学的には乳酸発酵とアルコール発酵に近い。 違いは、アルコール発酵の生成物がエチルアルコールと二酸化炭素であるということです。
ピルビン酸が二酸化炭素と水に酸化される次の段階は、 細胞呼吸。 呼吸関連の反応は、植物や動物の細胞のミトコンドリアで、酸素の存在下でのみ起こります。 これは、最終生成物である二酸化炭素が形成される前の一連の化学変換です。 このプロセスのさまざまな段階で、最初の物質の酸化の中間生成物が形成され、水素原子が除去されます。 この場合、ATPの化学結合に「保存」されているエネルギーが放出され、水分子が形成されます。 酸素が必要なのは、分離した水素原子を結合するためだけであることが明らかになります。 この一連の化学変換は非常に複雑であり、ミトコンドリア、酵素、および担体タンパク質の内膜の関与によって発生します。
細胞呼吸は非常に高い効率を持っています。 30個のATP分子の合成があり、解糖中にさらに2個の分子が形成され、ミトコンドリア膜での解糖生成物の変換の結果として6個のATP分子が形成されます。 合計で、1つのグルコース分子の酸化の結果として、38個のATP分子が形成されます。
C 6 H 12 O 6 + 6H20→6CO2+ 6H 2 O + 38ATP
ミトコンドリアでは、糖だけでなくタンパク質や脂質の酸化の最終段階が起こります。 これらの物質は、主に炭水化物の供給が終了したときに細胞によって使用されます。 まず、脂肪が消費され、その酸化中に、同量の炭水化物やタンパク質からよりもはるかに多くのエネルギーが放出されます。 したがって、動物の脂肪はエネルギー資源の主な「戦略的備蓄」です。 植物では、でんぷんはエネルギーの蓄えの役割を果たします。 保管すると、エネルギーに相当する量の脂肪よりもはるかに多くのスペースを占有します。 植物の場合、動物のように動かず、自分自身に蓄えを持たないので、これは障害ではありません。 あなたは脂肪からよりもはるかに速く炭水化物からエネルギーを抽出することができます。 タンパク質は体内で多くの重要な機能を果たします。したがって、タンパク質は、たとえば長期の飢餓時に糖や脂肪の資源が使い果たされた場合にのみ、エネルギー代謝に関与します。
光合成. 光合成-は、太陽光のエネルギーが有機化合物の化学結合のエネルギーに変換されるプロセスです。 で 植物細胞光合成に関連するプロセスは葉緑体で起こります。 このオルガネラの中には、太陽の放射エネルギーを取り込む色素が埋め込まれた膜のシステムがあります。 光合成の主な色素はクロロフィルで、主に青と紫、そしてスペクトルの赤い光線を吸収します。 緑の光同時に、それは反射されるので、クロロフィル自体とそれを含む植物の部分は緑色に見えます。
光合成には2つの段階があります- ライトと 暗い(図53)。 放射エネルギーの実際の捕捉と変換は、明期に行われます。 光量子を吸収すると、クロロフィルは励起状態になり、電子供与体になります。 その電子は、電子伝達系に沿って、あるタンパク質複合体から別のタンパク質複合体に移動します。 この鎖のタンパク質は、色素のように、葉緑体の内膜に集中しています。 電子がキャリアチェーンを通過すると、ATPの合成に使用されるエネルギーが失われます。 光によって励起された電子の一部は、NDP(ニコチンアミドアデニンジヌクレオチホスフェート)またはNADPHを還元するために使用されます。

米。 53.光合成の明期と暗期の反応の生成物
葉緑体の日光の影響下で、水分子の分裂も起こります- 光分解; この場合、クロロフィルによる損失を補う電子が発生します。 酸素は副産物として形成されます:
したがって、光相の機能的意味は、光エネルギーを化学エネルギーに変換することによるATPとNADP・Hの合成です。
光合成の暗期は光を必要としません。 ここで行われるプロセスの本質は、軽相で得られたATPおよびNADP・H分子が、炭水化物の形でCO2を「固定」する一連の化学反応で使用されることです。 暗相のすべての反応は葉緑体内で行われ、二酸化炭素の「固定」中に放出されたADPとNADPは、ATPとNADPHの合成のための明相の反応で再び使用されます。
全体的な光合成方程式は次のとおりです。
プラスチックとエネルギーの交換のプロセスの関係と統一。 ATP合成のプロセスは、細胞質(解糖)、ミトコンドリア(細胞呼吸)、および葉緑体(光合成)で発生します。 これらのプロセス中に発生するすべての反応は、エネルギー交換の反応です。 ATPの形で蓄えられたエネルギーは、細胞の寿命に必要なタンパク質、脂肪、炭水化物、および核酸の生産のためのプラスチック交換の反応に費やされます。 光合成の暗期は一連の反応、塑性交換であり、明期はエネルギーであることに注意してください。
エネルギーとプラスチック交換のプロセスの関係と統一性は、次の方程式でよく表されます。

この方程式を左から右に読むと、ATP(エネルギー代謝)の合成に関連して、解糖と細胞呼吸の間にグルコースを二酸化炭素と水に酸化するプロセスがわかります。 右から左に読むと、ATP(プラスチック代謝)が関与して水と二酸化炭素からブドウ糖が合成されるときの光合成の暗期の反応の説明が得られます。
化学合成。 光合成独立栄養菌に加えて、特定のバクテリア(水素、硝化、硫黄バクテリアなど)も無機物質から有機物質を合成することができます。 彼らは、無機物質の酸化中に放出されるエネルギーのためにこの合成を実行します。 それらは化学合成独立栄養体と呼ばれます。 これらの化学合成細菌は、生物圏で重要な役割を果たしています。 たとえば、硝化細菌は、植物がアクセスできないアンモニウム塩を硝酸塩に変換します。硝酸塩は、植物によく吸収されます。
細胞代謝は、エネルギーと塑性代謝の反応で構成されています。 エネルギー代謝の過程で、マクロ作動性の化学結合を持つ有機化合物(ATP)の形成が起こります。 これに必要なエネルギーは、嫌気的(解糖、発酵)および好気的(細胞呼吸)反応中の有機化合物の酸化から生じます。 太陽の光線から、そのエネルギーは光相に吸収されます(光合成); 無機化合物の酸化(化学合成)から。 ATPのエネルギーは、光合成の暗期の反応を含むプラスチック交換反応の過程で、細胞に必要な有機化合物の合成に費やされます。
- プラスチックとエネルギー代謝の違いは何ですか?
- 太陽光のエネルギーはどのようにして光合成の光相に変換されますか? 光合成の暗期にはどのようなプロセスが起こりますか?
- なぜ光合成は惑星と宇宙の相互作用の反射のプロセスと呼ばれるのですか?
エネルギーはATPの形で貯蔵され、ATPは物質の合成、熱の放出、 筋肉の収縮等
乳酸(筋肉に蓄積すると痛みを引き起こす可能性があります)は血液によって肝臓に運ばれ、そこで糖新生中にブドウ糖に変換されます。
アルコールは、アルコール発酵中に酵母細胞で形成されます。
アセチルCoA-脂肪酸、ケトン体、コレステロールなどの合成に使用されるか、クレブス回路で酸化されます。
水と二酸化炭素は一般的な代謝に含まれるか、体から排泄されます。
ペントースは、核酸、ブドウ糖(糖新生)、その他の物質の合成に使用されます。
NADPH2は、脂肪酸、プリン塩基などの合成に関与しています。 または、CPEでエネルギーを生成するために使用されます。
体内のブドウ糖の変換は、さまざまな酵素の作用下で発生するかなり複雑なプロセスです。 したがって、ブドウ糖から乳酸への経路には11の化学反応が含まれ、それぞれが独自の酵素によって加速されます。
スキーム番号8。 嫌気的解糖。
グルコース
ADPヘキソキナーゼ、Mgイオン
グルコース-6-リン酸
ホスホグルコイソメラーゼ
フルクトース6-リン酸
ADPホスホフルクトキナーゼ、Mgイオン
フルクトース1,6-二リン酸
アルドラーゼ
3-ホスホジオキシアセトン3-ホスホグリセロアルデヒド(3-PHA)
NADH +H3-PHAデヒドロゲナーゼ
1,3-ジホスホグリセリン酸
ATPホスホグリセリン酸ムターゼ
2-ホスホグリセリン酸
H2Oエノラーゼ
ホスホエノールピルビン酸
ATPピルビン酸キナーゼ、Mgイオン
ピルビン酸PVC
NAD乳酸デヒドロゲナーゼ
乳酸。
解糖は細胞の細胞質で起こり、ミトコンドリアの呼吸鎖を必要としません。
ブドウ糖は、すべての臓器や組織の細胞、特に神経系、赤血球、腎臓、精巣の主要なエネルギー源の1つです。
脳は、ほぼ完全に、拡散して入ってくるブドウ糖tkによって提供されます。 IVHは脳細胞に入りません。 したがって、血中のブドウ糖の濃度が低下すると、脳の機能が破壊されます。
糖新生。
嫌気性条件下では、ブドウ糖は骨格筋の働きのための唯一のエネルギー源です。 ブドウ糖から形成された乳酸は、次に血液に入り、肝臓に行き、そこでブドウ糖に変換され、次に筋肉に戻ります(コリ回路)。
非炭水化物物質をブドウ糖に変換するプロセスは、 糖新生。
糖新生の生物学的意義は次のとおりです。
飢餓や糖尿病など、体内に炭水化物が不足している場合に、ブドウ糖の濃度を十分なレベルに維持します。
乳酸、ピルビン酸、グリセロール、グリコーゲンアミノ酸、クレブス回路のほとんどの中間代謝物からのグルコースの形成。
糖新生は主に肝臓と腎皮質で起こります。 筋肉では、必要な酵素が不足しているため、このプロセスは発生しません。
糖新生の全反応:
2PVC + 4ATP + 2GTP + 2NADH + H + 4H2O
グルコース+2NAD+ 4ADP + 2GDP + 6H3PO4
したがって、糖新生の過程で、最大6つのマクロ作動性化合物と2NADH+Hが各グルコース分子に対して消費されます。
大量のアルコールを摂取すると糖新生が阻害され、脳機能の低下につながる可能性があります。 糖新生の速度は、以下の条件で増加する可能性があります。
断食するとき。
強化されたタンパク質栄養。
食品中の炭水化物の不足。
糖尿病。
グルコース代謝のグルクロン酸経路。
この経路は量的には重要ではありませんが、中和機能にとって非常に重要です。グルクロン酸の形で活性型のグルクロン酸(UDP-グルクロン酸)に結合する代謝最終産物と異物は、体から簡単に排泄されます。 グルクロン酸自体は、グリコサミノグリカンの必要な成分であるヒアルロン酸、ヘパリンなどです。ヒトでは、このグルコース分解経路の結果として、UDP-グルクロン酸が形成されます。



